
АНАТОМИЯ И ФИЗИОЛОГИЯ
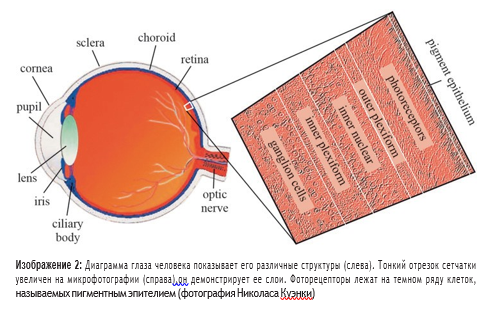
Понимание анатомии сетчатки приматов необходимо для понимания ее функций. У приматов фоторецепторы находятся в слое у задней части глазного яблока. Во втором из трех клеточных слоев, inner nuclear layer (INL), находятся от одного до четырех типов горизонтальных клеток, 11 типов биполярных клеток и от 22 до 30 типов амакринных клеток. Количества типов клеток варьируется в зависимости от вида животного. Поверхностный слой сетчатки содержит около 20 типов ганглиозных клеток. Информация от ганглиозных клеток поступает в мозг через более чем миллион волокон зрительного нерва. Пространства, разделяющие эти три слоя, также анатомически различны. Область, содержащая синапсы, связывающие фоторецепторы с биполярными и горизонтальными клетками, известна как outer plexiform layer (OPL), а область, где биполярные и амакриновые клетки соединяются с ганглиозными клетками, называется inner plexiform layer (IPL). Десятилетия анатомических исследований пролили свет на то, как работает сетчатка. Один из методов визуализации — окрашивание серебром, впервые применен Ramón y Cajal, а более ста лет назад был использован Гольджи. Сегодня исследователи применяют электронный микроскоп и окрашивание антител, что позволяет выявить формы и размеры типов клеток сетчатки и то, как различные клетки соединяются, образуя синапсы. Методы окрашивания выявили электрические соединения между клетками, а также идентичность и расположение рецепторов и транспортеров нейротрансмиттеров. Теперь мы знаем, что нейротрансмиттер, проходящий через вертикальные пути сетчатки — от фоторецепторов до биполярных клеток к ганглиозным клеткам — является глутаматом. Горизонтальные и амакринные клетки посылают сигналы с использованием различных возбуждающих и ингибирующих аминокислот, катехоламинов, пептидов и оксида азота. Электрофизиологические исследования сетчатки начались 60 лет назад. Исследования волокон зрительного нерва показали, что их можно стимулировать, чтобы создать традиционные потенциалы деполяризующего действия, аналогичные тем, которые наблюдаются в других нейронах. Однако первые записи импульсов в сетчатке, сделанные Gunnar Svaetichin в 1950-х годах, показали очень странные реакции на свет. Нейроны на внешней сетчатке (изначально не было понятно, какие именно) реагировали на стимуляцию не деполяризией, а медленной гиперполяризацией. В настоящее время известно, что эти «S-потенциалы» происходят от фоторецепторов и передают стимулируют биполярные клетки через горизонтальные. Гиперполяризация мембраны начинается при воздействии света и затем возвращается к исходному значению при отсутствии света. Это отражает противоречивый факт, что как палочки, так и колбочки высвобождают нейротрансмиттеры в темноте, когда мембрана деполяризована и ионы натрия свободно протекают через клеточные мембраны фоторецепторов. При воздействии света ионные каналы в клеточных мембранах закрываются. Клетки переходят в гиперполяризованное состояние до тех пор, пока свет продолжает попадать на них и в это время не высвобождают нейротрансмиттер. Хотя и палочки и колбочки реагируют на свет медленным гиперполяризационным откликом, они сообщают о совершенно разных свойствах ретинального изображения. Палочки, обнаруживающие тусклый свет, обычно реагируют на относительно медленные изменения. Колбочки действуют при ярком освещении, могут обнаруживать быстрые световые колебания. В обоих случаях фоторецепторы начинают процесс анализа изображений на отдельные части.И палочки, и колбочки реагируют на свет, падающий именно на них. Таким образом, их рецептивные поля очень узки. Изображение продолжает разбиваться на составляющие элементы в первых синапсах зрительного пути, между фоторецепторами и биполярными клетками. Различные биполярные клетки имеют разные типы рецепторов для нейротрансмиттера глутамата, что позволяет клеткам поразному реагировать на сигналы фоторецепторов (рис. 3). Некоторые биполярные клетки настроены на быстрые, а некоторые — на более медленные колебания визуального сигнала; некоторые рецепторы глутамата быстро повышают чувствительность, а другие более постепенно. Таким образом, клетки срабатывают либо быстро по очереди, либо относительно медленно в ответ на одинаковое количество стимуляции. Эти рецепторы реагируют на глутамат, активируя так называемый путь ВЫКЛ (OFF pathway) в визуальном процессе, обнаруживая темные изображения на более светлом фоне. (Напомним, что фоторецепторы постоянно высвобождают глутамат, если не подвергаются воздействию света.) Другие биполярные клетки имеют ингибирующие рецепторы глутамата; иными словами, они предотвращают срабатывание биполярной клетки, когда она подвергается воздействию нейротрансмиттера. Эти рецепторы активируют путь ВКЛ (ON pathway), обнаруживая светлые изображения на темном фоне.
Наталья Ринская

")
")

")
")
")
")
")

")
")

")

")
")




")

