ПОСТОРОННИЕ ИЗОБРАЖЕНИЯ ПРИ ПОМОЩИ АМАКРИНОВЫХ КЛЕТОК СЕТЧАТКИ
Обработка информации происходит еще до того, как ганглиозные клетки передают сигнал в мозг. Для этого важно знать организацию IPL, где 22 или более различных типов амакринных клеток имеют синаптические связи с примерно 20 различными типами ганглиозных клеток. Из описания Кахаля в 19 веке уже было ясно, что амакринные, ганглиозные и биполярноклеточные дендриты и аксоны организованы в отдельные слои. Сам Кахал разделил внутренний плексиформный слой на пять слоев. Но какие синапсы были сформированы среди путаницы взаимосвязанных процессов и как именно все организованно, не было сразу очевидно. Электронная микроскопия помогла распутать эту нейроциркуляцию. В настоящее время взаимосвязь девяти типов биполярных клеток, 14 типов амакринных клеток и восьми типов ганглиозных клеток изучена достаточно хорошо. Можно сказать, что мы находимся на полпути к пониманию нейронного взаимодействия между всеми нервными клетками сетчатки. В настоящее время известно много о том, какие типы нейротрансмиттеров содержатся в разных амакринных клетках, а также об организации рецепторов в разных синапсах. Амакринные клетки примерно поровну поделены между теми, которые используют нейромедиаторы глицина, и теми, которые используют ГАМК (гамма-аминомасляную кислоту). Глицинергические амакриновые клетки обычно имеют «маленькие поля». Их поле обработки информации может распространяться вертикально через несколько слоев в IPL, но оно распространяется относительно на короткие расстояния горизонтально. Глицинергические амакриновые клетки получают информацию от биполярных клеток и передают информацию ганглиозным клеткам и другим биполярным и амакринным клеткам. Некоторые глицинергические амакриновые клетки обеспечивают взаимосвязь между системами ON и OFF биполярных и ганглиозных клеток. В более раннем обсуждении каналов ON и OFF, исходящих из колбочек, не обсуждался канал из палочек. Колбочки соединяются напрямую с биполярными клетками, передавая сигнал ганглиозным клеткам, а биполярные клетки, которые получают ввод от палочек, не имеют синапса с ганглиозными клетками напрямую. Все биполярные клетки, соединенные с палочками, относятся к одному типу и передают только сигнал ON и соединяются с амакриновыми AII cell и A17, которые используют в качестве посредников для получения сигналов для ганглиозных клеток. В то же время амакриновая клетка A17 собирает сообщения палочек из тысяч палочковых биполярных клеток. Она каким-то образом усиливает и модулирует информацию от палочковых биполярных клеток для ее передачи в AII cells, но как это происходит не совсем понятно. В любом случае, информационный путь палочек с его серией сходящихся и затем расходящихся промежуточных нейронов, явно хорошо спроектирован для сбора и усиления рассеянных остатков света в условиях сумеречного и ночного зрения. Амакринные клетки с широкими рецептивными полями иногда растягиваются горизонтально через IPL на сотни микрон и взаимодействуют с сотнями биполярных клеток и многими ганглиозными клетками. Такие амакриновые клетки принадлежат к одной из пяти различных страт внутреннего плексиформного слоя и создают изящные сетки дендритов. Обычно они высвобождают ГАМК в качестве нейротрансмиттера. Иногда они соединяются с соседними амакринными клетками с помощью щелевых контактов, увеличивая сферу своего влияния и скорость, с которой сигналы передаются через большие области сетчатки. Большинство амакриновых клеток, высвобождающих ГАМК, также выделяют по меньшей мере одно дополнительное нейроактивное вещество. Вторичные вещества обычно являются нейромодуляторами, а не быстродействующими нейротрансмиттерами. К этим веществам относятся пептиды — «вещество Р», соматостатин, вазоинтестинальный пептид и холецистокинин, а также более известные биомолекулы — серотонин, дофамин, ацетилхолин, аденозин и оксид азота. На ганглиозных и биполярных клетках было обнаружено множество рецепторов, например, рецепторы для пептидов, никотина и мускарина (грибной токсин) в дополнение к различным формам ГАМК-рецепторов, что указывает на то, что амакриновые клетки высвобождают такие агенты. Большинство из этих нейромодуляторов не активны в обычных синапсах; считается, что их высвобождение влияет на нейроны даже на расстоянии путем диффузии. Такие нейромодуляторы, по-видимому, влияют на схему сетчатки при изменении условий освещения или даже заставляют сетчатку отражать различное время дня в циркадных ритмах. Специализированная амакриновая клетка высвобождает дофамин когда сетчатка стимулируется прерывистым мигающим светом. Дофамин разъединяет щелевые соединения между горизонтальными клетками, уменьшая размер их рецептивных полей. Кроме того, нейромедиатор воздействует на рецептор глутамата на горизонтальных клетках, так что амплитуда светового ответа снижается. Снова в IPL допамин закрывает щелевые соединения, на этот раз те, которые связывают AII cells в больших нейронных сетях. В результате разобщение AII ceels делает эффективное рецептивное поле систем палочковых амакринных клеток гораздо менее значимым в условиях яркого освещения. 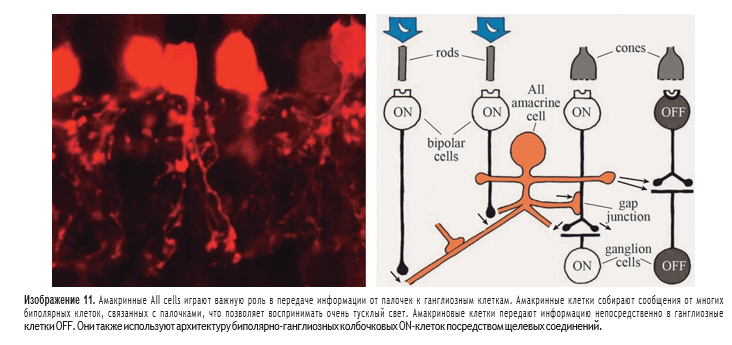 Точно так же при ярком освещении другая амакриновая клетка с широким рецептивным полем высвобождает оксид азота, чтобы отделить AII cell от колбочковой-биполярной системы. Все это устраняет вмешательство палочек с широким рецептивным полем в узкое рецептивное поле колбочек, которое обеспечивает колбочковый зрительный путь. Приведенная выше схема сетчатки позволяет предположить, что сетчатка удивительно сложна. По мере развития исследований становится ясно, что сетчатка играет активную роль в зрительном восприятии. Хотя мы не полностью понимаем нейронный код, который аксоны ганглиозных клеток посылают в мозг, мы приближаемся к пониманию того, как ансамбли ганглиозных клеток по-разному реагируют на аспекты зрительного восприятия окружающего мира и как строятся поля влияния на отдельные ганглиозные клетки. Значимая часть восприятия зрительного объекта, происходит в самой сетчатке, хотя окончательное восприятие бесспорно осуществляется в головном мозге. Не так давно было обнаружено, что большая часть переноса информации зависит от электрических связей между клетками, а не от химических синапсов. Например, основной неврологический путь от палочек зависит от прямых электрических соединений. Некоторые другие быстродействующие сигналы передаются от амакринных клеток в ганглиозные клетки в щелевых соединениях. Нейромодуляторы изменяют среду нейронных контуров, но действуют на отдаленном расстояния посредством диффузии, а не в близко расположенных синапсах. Опять же, это удивительная концепция по сравнению с предыдущим представлением о том, что все нейронные взаимодействия происходят через нейротрансмиттеры в специфически изолированных участках мембран, то есть синапсах. H. Kolb 2003
Точно так же при ярком освещении другая амакриновая клетка с широким рецептивным полем высвобождает оксид азота, чтобы отделить AII cell от колбочковой-биполярной системы. Все это устраняет вмешательство палочек с широким рецептивным полем в узкое рецептивное поле колбочек, которое обеспечивает колбочковый зрительный путь. Приведенная выше схема сетчатки позволяет предположить, что сетчатка удивительно сложна. По мере развития исследований становится ясно, что сетчатка играет активную роль в зрительном восприятии. Хотя мы не полностью понимаем нейронный код, который аксоны ганглиозных клеток посылают в мозг, мы приближаемся к пониманию того, как ансамбли ганглиозных клеток по-разному реагируют на аспекты зрительного восприятия окружающего мира и как строятся поля влияния на отдельные ганглиозные клетки. Значимая часть восприятия зрительного объекта, происходит в самой сетчатке, хотя окончательное восприятие бесспорно осуществляется в головном мозге. Не так давно было обнаружено, что большая часть переноса информации зависит от электрических связей между клетками, а не от химических синапсов. Например, основной неврологический путь от палочек зависит от прямых электрических соединений. Некоторые другие быстродействующие сигналы передаются от амакринных клеток в ганглиозные клетки в щелевых соединениях. Нейромодуляторы изменяют среду нейронных контуров, но действуют на отдаленном расстояния посредством диффузии, а не в близко расположенных синапсах. Опять же, это удивительная концепция по сравнению с предыдущим представлением о том, что все нейронные взаимодействия происходят через нейротрансмиттеры в специфически изолированных участках мембран, то есть синапсах. H. Kolb 2003 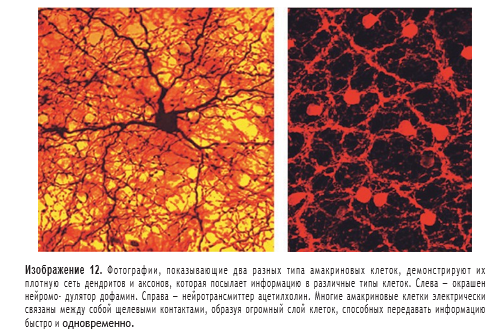
Наталья Ринская

")
")

")
")
")
")
")

")
")

")

")
")




")

